Termini e concetti chiave della genetica
Genetica e Ereditarietà
Lo studio dei fenomeni ereditari in biologia è compito della genetica, l'insieme dei processi e delle modalità relativi alla trasmissione dei caratteri genetici da un organismo alla sua progenie e, quindi, da una generazione alla successiva; è detta ereditarietà
In forma riassuntiva, è la proprietà caratteristica degli esseri viventi di trasmettere alla progenie, per mezzo del patrimonio genetico, le informazioni relative ai caratteri morfologici e fisiologici (o patrimonio ereditario)
Poligenesi: origine molteplice, derivazione da ceppi diversi (direttamente contrapposto a monogenesi). In biologia, il termine indica l’origine di un gruppo di organismi da forme primitive diverse, anziché da una sola (➔ Monogenesi), o l’origine di un nuovo gruppo in più località o più volte nel tempo.
Cellula: il corpo di tutti gli organismi è composto da organi (epiteli, muscoli, scheletro, apparato digerente, apparato respiratorio, ecc...) ognuno dei quali è formato da miliardi di cellule. La cellula rappresenta quindi il 'mattone' fondamentale con cui costruire forme di vita complessa. Visto che in genere un corpo usa nascere piccolo e poi crescere, è naturale che, non potendo le cellule aumentare di dimensioni, debbano necessariamente aumentare numericamente. Questo processo implica però un passo fondamentale ovvero ogni cellula deve continuare ad avere tutta l'informazione necessaria per compiere il proprio compito all'interno dell'organismo.
Nucleo cellulare e DNA: L'informazione necessaria si trova nel nucleo: un apposito compartimento cellulare che separa il DNA dal resto della cellula (citoplasma, reticolo endoplasmico, mitocondri ecc).
L'acido desossiribonucleico o deossiribonucleico (DNA) è un acido nucleico che contiene le informazioni genetiche necessarie alla biosintesi di RNA e proteine, le molecole indispensabili per lo sviluppo ed il corretto funzionamento della maggior parte degli organismi viventi.
Poligenesi: origine molteplice, derivazione da ceppi diversi (direttamente contrapposto a monogenesi). In biologia, il termine indica l’origine di un gruppo di organismi da forme primitive diverse, anziché da una sola (➔ Monogenesi), o l’origine di un nuovo gruppo in più località o più volte nel tempo.
Penetranza = capacità di un allele dominante di manifestarsi fenotipicamente:
_ si valuta su di una popolazione, non su un singolo individuo, in proporzione quindi agli individui che manifestano il carattere rispetto al totale che portano quell’allele.
Un carattere che si manifesta nell’80% della popolazione vuol dire che ha una penetranza dell’80%
Penetranza incompleta = la non espressione del carattere negli individui che portano il relativo allele
Espressività = intensità con cui un gene si manifesta a livello fenotipico e può variare da elevata a nulla (in assenza di penetranza del gene), il concetto di espressività non si intende a livello del singolo individuo ma in termini quantitativi
Polimorfismo: presenza in una popolazione di due o più genotipi per un dato carattere, o anche l’esistenza di una variazione fenotipica in una popolazione.
DIVISIONE CELLULARE
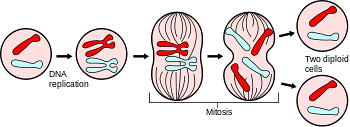
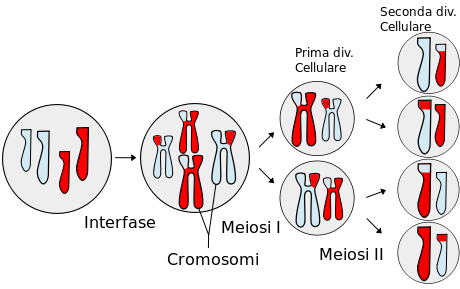 La meiosi è un processo di divisione mediante il quale una cellula eucariotica con corredo cromosomico diploide dà origine a quattro cellule con corredo cromosomico aploide, da una cellula madre si formano quattro cellule figlie, tutte diverse fra loro.
La meiosi è un processo di divisione mediante il quale una cellula eucariotica con corredo cromosomico diploide dà origine a quattro cellule con corredo cromosomico aploide, da una cellula madre si formano quattro cellule figlie, tutte diverse fra loro.
Potrebbe sembrare molto simile alla mitosi ma, al contrario di questa, si ha la riduzione da corredo in doppia copia a corredo a semplice copia, e tramite il cosiddetto crossing-over (incrocio esterno), si ha lo scambio e la ricombinazione genetica, che poi sta alla base dell'evoluzione.
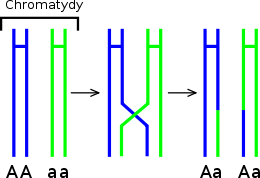
Il crossing-over è l'importante meccanismo di ricombinazione del materiale genetico proveniente dai due genitori, che permette una maggiore varietà nei prodotti della riproduzione sessuata.
Dominanza: mutazione genetica espressa sia in eterozigosi che in omozigosi.
In genetica, si parla di dominanza incompleta quando un alelle è dominante sull'altro, ma non in modo completo. Ne consegue che l'altro allele ha possibilità di esprimersi, anche se in misura minore rispetto all'allele dominante. Il fenotipo manifestato dall'eterozigote è un fenotipo intermedio tra quelli dei due omozigoti (dominante e recessivo).
Differenza dalla codominanza:
_ nella codominanza gli alleli di un medesimo gene sono espressi con piena funzionalità contemporaneamente, come accade nel gruppo sanguigno AB, in cui sono espressi efficacemente entrambi i geni per gli antigeni A e B.
_ nella dominanza incompleta, invece, accade non solo che nell'eterozigote, come avviene anche nella dominanza completa, l'allele mutato è o amorfo, cioè inattivo, o ipomorfo, cioè con attività qualitativamente normale ma quantitativamente ridotta, ma contemporaneamente si osserva anche e soprattutto un'aploinsufficienza della copia selvatica del gene, cioè l'incapacità di una sola copia selvatica del gene per cellula a garantirne una funzione normale, perché per un fenotipo penetrante è richiesto un dosaggio molecolare del prodotto genico più elevato che in eterozigosi.
Il BOA PASTEL
 (Sopra Bci W.Pastel)
(Sopra Bci W.Pastel)

(Sopra: Bci Red Pastel eccellente)

(Sopra: Bci HiperPastel = doppio Pastel eccellente x doppio Pastel eccellente)
I Boa presentati in queste foto sono un'eccellenza selettiva ottenuta prelevando da molteplici cucciolate gli esemplari dalle caratteristiche cromatiche di spicco più estreme ricombinate assieme per più generazioni.

Per via del loro magnifico patrimonio genetico i veri Pastel accoppiati con altri Boa comuni o morph sono in grado di trasmettere le loro importanti caratteristiche di colorazione e pulizia alla progene diminuendo le antiestetiche puntinature che ne "sporcherebbero" il fondo, migliorandone così l'aspetto.
Solo i ceppi Pastel eccellenti infatti possono essere utilizzati con successo per migliorare e valorizzare ulteriori selezioni genetiche di Boa morph,..
.. vedi come esempio il Pastel Ivory (foto sopra) utilizzato con il morph Anery (foto sotto).
(Sotto: Bci Anery Pastel Ivory)

_____________________________________________________________________R.R.__
BIOGRAFIE:
 IL PADRE DELLA GENETICA
IL PADRE DELLA GENETICA
È stato solo dopo il 1900, successivamente alla riscoperta delle sue Leggi, che sono stati compresi i suoi risultati sperimentali.
 REGINALD CRUNDALL PUNNETT (1875-1967)
REGINALD CRUNDALL PUNNETT (1875-1967)
Andò alla Cambridge University come studente di medicina, Punnett ma ottenne laurea in zoologia nel 1898. Dopo la laurea continuò come ricercatore a Cambridge. Egli lavorò sulla morfologia dei Nemertini (vermi piatti). A Punnett sono state dedicate due specie di vermi marini: Cerbratulus punnetti, Punnettia splendia.


 Carl Nilsson Linnaeus, divenuto Carl von Linné in seguito all'acquisizione di un titolo nobiliare e noto ai più semplicemente come Linneo (dalla forma latinizzata del nome, Carolus Linnaeus) (Råshult, 23 maggio 1707 – Uppsala, 10 gennaio 1778), è stato un medico e naturalista svedese, considerato il padre della moderna classificazione scientifica degli organismi viventi.
Carl Nilsson Linnaeus, divenuto Carl von Linné in seguito all'acquisizione di un titolo nobiliare e noto ai più semplicemente come Linneo (dalla forma latinizzata del nome, Carolus Linnaeus) (Råshult, 23 maggio 1707 – Uppsala, 10 gennaio 1778), è stato un medico e naturalista svedese, considerato il padre della moderna classificazione scientifica degli organismi viventi.
La lettera L., posta spesso a seguire delle indicazioni di nomenclatura binomiale nei cataloghi di specie, identifica il cognome dello scienziato.,Il merito maggiore dello svedese fu la definizione e l'introduzione nel 1735 della nomenclatura binomiale, basata sul modello aristotelico di definizione mediante genere prossimo e differenza specifica, nel sistema di classificazione delle piante e degli animali.[2] Con questo metodo tassonomico (concepito poco più di un secolo prima da Bauhin) a ciascun organismo sono attribuiti due nomi (di origine latina): il primo si riferisce al Genere di appartenenza dell'organismo stesso ed è uguale per tutte le specie che condividono alcuni caratteri principali (nomen genericum); il secondo termine, che è spesso descrittivo, designa la Specie propriamente detta (nome triviale o nome specifico).
La portata dell’innovazione fu enorme; precedentemente alla nomenclatura binomiale il sistema di nomenclatura era semplicemente basato su un'estesa descrizione di ogni pianta, in latino, per i caratteri distintivi ritenuti di rilievo, in modo del tutto arbitrario, da ogni classificatore.

 http://www.associazionelinnaeus.it/
http://www.associazionelinnaeus.it/

In forma riassuntiva, è la proprietà caratteristica degli esseri viventi di trasmettere alla progenie, per mezzo del patrimonio genetico, le informazioni relative ai caratteri morfologici e fisiologici (o patrimonio ereditario)
Le leggi dell’ereditarietà: i meccanismi fondamentali dell’ereditarietà furono identificati da G. Mendel che formulò le tre leggi (dette della dominanza, della segregazione e dell’assortimento indipendente dei caratteri), tuttora alla base della genetica classica.
Mendel per primo postulò la natura particolata delle unità ereditarie e introdusse il concetto di allele, di dominanza e di recessività dei caratteri.
Oggi le modalità della trasmissione dei caratteri sono state spiegate a livello molecolare; si conosce non solo la localizzazione fisica (locus genico; plurale "loci") delle unità ereditarie (i geni), ma anche la loro composizione biochimica e la loro sequenza nucleotidica.
Ogni organismo appartenente alle specie che si riproducono sessualmente eredita due copie di ciascun gene, dette alleli, ciascuna proveniente da uno dei due genitori. Un individuo, i cui due alleli determinanti un carattere sono uguali, si dice omozigote, se diversi tra loro, si dice eterozigote. Quando in un individuo si originano le cellule sessuali, ciascuno dei risultanti gameti riceve un membro di ciascuna coppia allelica.
Ogni genitore trasmette metà dei suoi geni alla generazione successiva, e figli degli stessi genitori ricevono differenti combinazioni di geni, grazie al fenomeno della ricombinazione.
Costanza e variabilità nella specie.
Il concetto di ereditarietà comprende due aspetti apparentemente contraddittori fra loro. Si può infatti osservare negli organismi viventi la costanza delle specie da una generazione all’altra, e la variabilità tra individui all’interno della stessa specie.
Costanza e variabilità della specie sono in realtà entrambi aspetti dell’ereditarietà che possono essere spiegati simultaneamente dalla funzione dei geni, le unità funzionali di materiale genetico che si trovano in tutte le cellule degli esseri viventi. Ogni membro di una specie ha un assetto di geni specifici che forniscono le caratteristiche di quella specie (➔ genoma), e le mantengono attraverso le generazioni.
Tuttavia, tra gli individui della stessa specie possono avvenire variazioni all’interno di ciascun gene, che sono alla base del fatto che nessun individuo ha esattamente gli stessi caratteri di un altro.
Il concetto di ereditarietà comprende due aspetti apparentemente contraddittori fra loro. Si può infatti osservare negli organismi viventi la costanza delle specie da una generazione all’altra, e la variabilità tra individui all’interno della stessa specie.
Costanza e variabilità della specie sono in realtà entrambi aspetti dell’ereditarietà che possono essere spiegati simultaneamente dalla funzione dei geni, le unità funzionali di materiale genetico che si trovano in tutte le cellule degli esseri viventi. Ogni membro di una specie ha un assetto di geni specifici che forniscono le caratteristiche di quella specie (➔ genoma), e le mantengono attraverso le generazioni.
Tuttavia, tra gli individui della stessa specie possono avvenire variazioni all’interno di ciascun gene, che sono alla base del fatto che nessun individuo ha esattamente gli stessi caratteri di un altro.
La variabilità tra organismi è, in generale, di natura quantitativa
Diversi fenotipi controllati da numerosi genotipi (all’aumentare del numero di loci che controllano un carattere), aumenta il numero di genotipi.
Si parla di Ereditabilità di un carattere solo se la somiglianza deriva dal condividere lo stesso genotipo.
La dimensione, il colore della pelle sono caratteri ereditabili, nel miglioramento genetico, la variabilità genetica può portare a miglioramenti nelle colorazioni,dimensioni e altre caratteristiche importanti.
Si parla di Ereditabilità di un carattere solo se la somiglianza deriva dal condividere lo stesso genotipo.
La dimensione, il colore della pelle sono caratteri ereditabili, nel miglioramento genetico, la variabilità genetica può portare a miglioramenti nelle colorazioni,dimensioni e altre caratteristiche importanti.
La maggior parte dei geni esiste in una popolazione sotto forma di numerose varianti alleliche, e molti caratteri sono poligenici (ossia influenzati da più di un gene).
La natura poligenica e la molteplicità di alleli di molti caratteri sono all’origine del vasto potenziale di variabilità fra le caratteristiche ereditarie.Caratteri codificati da più loci sono detti caratteri poligenici
Poligenesi: origine molteplice, derivazione da ceppi diversi (direttamente contrapposto a monogenesi). In biologia, il termine indica l’origine di un gruppo di organismi da forme primitive diverse, anziché da una sola (➔ Monogenesi), o l’origine di un nuovo gruppo in più località o più volte nel tempo.
Cellula: il corpo di tutti gli organismi è composto da organi (epiteli, muscoli, scheletro, apparato digerente, apparato respiratorio, ecc...) ognuno dei quali è formato da miliardi di cellule. La cellula rappresenta quindi il 'mattone' fondamentale con cui costruire forme di vita complessa. Visto che in genere un corpo usa nascere piccolo e poi crescere, è naturale che, non potendo le cellule aumentare di dimensioni, debbano necessariamente aumentare numericamente. Questo processo implica però un passo fondamentale ovvero ogni cellula deve continuare ad avere tutta l'informazione necessaria per compiere il proprio compito all'interno dell'organismo.
Nucleo cellulare e DNA: L'informazione necessaria si trova nel nucleo: un apposito compartimento cellulare che separa il DNA dal resto della cellula (citoplasma, reticolo endoplasmico, mitocondri ecc).
L'acido desossiribonucleico o deossiribonucleico (DNA) è un acido nucleico che contiene le informazioni genetiche necessarie alla biosintesi di RNA e proteine, le molecole indispensabili per lo sviluppo ed il corretto funzionamento della maggior parte degli organismi viventi.
Ogni volta che una cellula si divide deve necessariamente passare l'intera informazione ad ambedue le cellule 'figlie' quindi deve duplicare quel lungo filamento
Locus genico: in biologia e in computazione evoluzionistica, il termine locus genico (o più semplicemente locus, plurale loci) designa la posizione di un gene o di un'altra sequenza significativa all'interno di un cromosoma.Un locus può essere occupato da uno qualsiasi degli alleli del gene.
Le cellule diploidi o poliploidi possono essere definite omozigoti se possiedono il medesimo allele nello stesso locus in ogni cromosoma, o eterozigoti, se possiedono diversi alleli nei loci corrispondenti.
Una mappa genetica (prodotta utilizzando gli incroci genetici) indica proprio la localizzazione, o meglio il locus, di ciascun gene che è stato mappato).
Il cromosoma è l'unità strutturale in cui il DNA, associato con specifiche proteine, si organizza all'interno delle cellule. I cromosomi recano su di essi l'informazione genetica. Il termine "cromosoma" deriva dal greco croma che significa "colore", e soma che significa "corpo"
Il corredo cromosomico di un essere vivente può essere definito l'insieme dei cromosomi presenti in una cellula somatica, tranne quelle sessuali, ed è costante a livello di specie, sia sotto il profilo numerico sia sotto quello morfologico. Ogni variazione nel corredo cromosomico può portare a delle variazioni anche consistenti nel fenotipo dell'individuo che ne deriva queste differenze possono anche essere migliorative e vengono premiate dall'evoluzione, ma a volte anche negative potendo portare alla morte dell'individuo o a gravi malformazioni.
Il corredo cromosomico di una specie viene indicato con 2n perché in genere i cromosomi sono presenti in duplice copia, i cui due componenti (cromosomi analoghi) hanno uno origine materna e l'altro paterna.
Nei corredi cromosomici delle specie animali sono in genere presenti anche due cromosomi particolari (cromosomi sessuali) coinvolti nei meccanismi di determinazione del sesso.
Geni e alleli: i caratteri di un organismo vengono determinati dai geni, ognuno dei quali occupa un preciso locus sul DNA. Negli individui diploidi ogni singolo gene consiste in una coppia di alleli, di cui uno viene passato dal padre e uno dalla madre: la coppia di alleli determinerà il fenotipo dell'animale.
Locus genico: in biologia e in computazione evoluzionistica, il termine locus genico (o più semplicemente locus, plurale loci) designa la posizione di un gene o di un'altra sequenza significativa all'interno di un cromosoma.Un locus può essere occupato da uno qualsiasi degli alleli del gene.
Le cellule diploidi o poliploidi possono essere definite omozigoti se possiedono il medesimo allele nello stesso locus in ogni cromosoma, o eterozigoti, se possiedono diversi alleli nei loci corrispondenti.
Una mappa genetica (prodotta utilizzando gli incroci genetici) indica proprio la localizzazione, o meglio il locus, di ciascun gene che è stato mappato).
Il cromosoma è l'unità strutturale in cui il DNA, associato con specifiche proteine, si organizza all'interno delle cellule. I cromosomi recano su di essi l'informazione genetica. Il termine "cromosoma" deriva dal greco croma che significa "colore", e soma che significa "corpo"
Il corredo cromosomico di un essere vivente può essere definito l'insieme dei cromosomi presenti in una cellula somatica, tranne quelle sessuali, ed è costante a livello di specie, sia sotto il profilo numerico sia sotto quello morfologico. Ogni variazione nel corredo cromosomico può portare a delle variazioni anche consistenti nel fenotipo dell'individuo che ne deriva queste differenze possono anche essere migliorative e vengono premiate dall'evoluzione, ma a volte anche negative potendo portare alla morte dell'individuo o a gravi malformazioni.
Il corredo cromosomico di una specie viene indicato con 2n perché in genere i cromosomi sono presenti in duplice copia, i cui due componenti (cromosomi analoghi) hanno uno origine materna e l'altro paterna.
Nei corredi cromosomici delle specie animali sono in genere presenti anche due cromosomi particolari (cromosomi sessuali) coinvolti nei meccanismi di determinazione del sesso.
Esistono alleli dominanti e recessivi: i primi manifestano il loro carattere a prescindere dall’altro, i secondi manifestano la mutazione solo quando entrambi gli alleli sono uguali.
Da qui due concetti base per comprendere i meccanismi che stanno dietro alle mutazioni genetiche(morph):
_ in omozigosi entrambi gli alleli sono uguali mentre in eterozigosi gli alleli sono diversi.
Morph = Mutazione genetica dovuta dalla presenza di uno o più alleli mutati rispetto alla forma normale.
_ in omozigosi entrambi gli alleli sono uguali mentre in eterozigosi gli alleli sono diversi.
Morph = Mutazione genetica dovuta dalla presenza di uno o più alleli mutati rispetto alla forma normale.
Eterozigosi: i due alleli di una specifica coppia sono diversi (uno è mutato e l'altro no).
Omozigosi: i due alleli di una specifica coppia sono uguali (sono entrambi mutati o entrambi normali).
Genotipo: profilo e costituzione genetica di un individuo, ossia il patrimonio ereditario di un individuo che compone la totalità dei suoi geni nel DNA.
Fenotipo: in relazione al genotipo indica sia la manifestazione esteriore(aspetto) di un tratto somatico nella specifica espressione di un gene, che l’insieme dei caratteri ereditari morfologici visibili.
Il set di geni che la prole eredita da entrambi i genitori, come combinazione del materiale genetico di ciascuno di essi, viene chiamato genotipo.
Il fenotipo (ossia le caratteristiche osservabili di una cellula o di un organismo), è invece il risultato dell’espressione genica (➔), comprende la struttura fisica di un organismo, i processi fisiologici e il comportamento.
Sebbene il genotipo determini i limiti delle caratteristiche che un organismo può sviluppare, il fenotipo dipende da interazioni complesse fra i geni e il loro ambiente.
Il genotipo rimane costante attraverso tutta la vita dell’organismo, invece, poiché l’ambiente interno e esterno cambiano continuamente, anche il fenotipo cambia.
Omozigosi: i due alleli di una specifica coppia sono uguali (sono entrambi mutati o entrambi normali).
Genotipo: profilo e costituzione genetica di un individuo, ossia il patrimonio ereditario di un individuo che compone la totalità dei suoi geni nel DNA.
Fenotipo: in relazione al genotipo indica sia la manifestazione esteriore(aspetto) di un tratto somatico nella specifica espressione di un gene, che l’insieme dei caratteri ereditari morfologici visibili.
Il set di geni che la prole eredita da entrambi i genitori, come combinazione del materiale genetico di ciascuno di essi, viene chiamato genotipo.
Il fenotipo (ossia le caratteristiche osservabili di una cellula o di un organismo), è invece il risultato dell’espressione genica (➔), comprende la struttura fisica di un organismo, i processi fisiologici e il comportamento.
Sebbene il genotipo determini i limiti delle caratteristiche che un organismo può sviluppare, il fenotipo dipende da interazioni complesse fra i geni e il loro ambiente.
Il genotipo rimane costante attraverso tutta la vita dell’organismo, invece, poiché l’ambiente interno e esterno cambiano continuamente, anche il fenotipo cambia.
Poligenesi: origine molteplice, derivazione da ceppi diversi (direttamente contrapposto a monogenesi). In biologia, il termine indica l’origine di un gruppo di organismi da forme primitive diverse, anziché da una sola (➔ Monogenesi), o l’origine di un nuovo gruppo in più località o più volte nel tempo.
Penetranza = capacità di un allele dominante di manifestarsi fenotipicamente:
_ si valuta su di una popolazione, non su un singolo individuo, in proporzione quindi agli individui che manifestano il carattere rispetto al totale che portano quell’allele.
Un carattere che si manifesta nell’80% della popolazione vuol dire che ha una penetranza dell’80%
Penetranza incompleta = la non espressione del carattere negli individui che portano il relativo allele
Evoluzione = variazione genetica nel tempo entro gruppi di organismi, il potenziale evolutivo immediato di una popolazione è determinato dall’ereditabilità dei suoi caratteri fenotipici
Selezione artificiale = mezzo per produrre un rapido cambiamento evolutivo, è competenza dell'allevatore che con cognizione acquisita dall'esperienza seleziona gli individui che dovranno riprodursi.
Se i caratteri selezionati avranno una base genetica, anch’essi varieranno e si evolveranno
Polimorfismo: presenza in una popolazione di due o più genotipi per un dato carattere, o anche l’esistenza di una variazione fenotipica in una popolazione.
DIVISIONE CELLULARE
La mitòsi è la riproduzione per divisione equazionale della cellula procariote . Il termine viene molto spesso utilizzato anche per la riproduzione delle cellule procariote, un processo molto più semplice e più correttamente chiamato scissione binaria o amitosi. Il termine mitosi deriva dal greco mìtos, "filo"; nome dovuto all'aspetto filiforme dei cromosomi durante la metafase
La mitosi è molto simile alla meiosi, si distinguono dal fatto che la mitosi forma 2 cellule diploidi con lo stesso numero di cromosomi della cellula madre, invece nella meiosi si formano 4 cellule aploidi con il corredo cromosomico dimezzato.
La mitosi riguarda le cellule somatiche dell'organismo (ossia tutte le cellule fuorché quelle che hanno funzione riproduttiva: i gametociti primari i quali vanno incontro alla meiosi e le cellule germinali ancora indifferenziate.
Potrebbe sembrare molto simile alla mitosi ma, al contrario di questa, si ha la riduzione da corredo in doppia copia a corredo a semplice copia, e tramite il cosiddetto crossing-over (incrocio esterno), si ha lo scambio e la ricombinazione genetica, che poi sta alla base dell'evoluzione.
La meiosi è fondamentale nella riproduzione sessuale, la ricombinazione dell'informazione genetica proveniente dalle cellule di due organismi differenti (uomo e donna) produce risultati ogni volta diversi, e naturalmente diversi anche dai due genitori.
Ogni genitore fornisce un corredo cromosomico "semplice" aploide (detto anche "dimezzato"), cellula uovo nella femmina e spermatozoo nel maschio; la fusione (fecondazione) dei due corredi dimezzati (materno e paterno) e "rimescolati" ricostituisce il corredo intero e dà origine ad una singola nuova cellula, detta zigote, che diverrà il nuovo individuo.
Ogni genitore fornisce un corredo cromosomico "semplice" aploide (detto anche "dimezzato"), cellula uovo nella femmina e spermatozoo nel maschio; la fusione (fecondazione) dei due corredi dimezzati (materno e paterno) e "rimescolati" ricostituisce il corredo intero e dà origine ad una singola nuova cellula, detta zigote, che diverrà il nuovo individuo.
Il crossing-over è l'importante meccanismo di ricombinazione del materiale genetico proveniente dai due genitori, che permette una maggiore varietà nei prodotti della riproduzione sessuata.
Dominanza: mutazione genetica espressa sia in eterozigosi che in omozigosi.
Un gene è definito dominante quando esprime fenotipicamente la sua mutazione sia in omozigosi sia in eterozigosi.
Codominanza e dominanza incompleta: la mutazione in omozigosi è diversa da quella in eterozigosi e recessiva quando la mutazione si manifesta solo ed unicamente in omozigosi.In genetica, si parla di dominanza incompleta quando un alelle è dominante sull'altro, ma non in modo completo. Ne consegue che l'altro allele ha possibilità di esprimersi, anche se in misura minore rispetto all'allele dominante. Il fenotipo manifestato dall'eterozigote è un fenotipo intermedio tra quelli dei due omozigoti (dominante e recessivo).
Per esempio poniamo di essere di fronte a una popolazione la cui sopravvivenza sia dovuta in gran parte alla capacità dell'individuo di mimetizzarsi. Poniamo di trovare al locus del mimetismo due alleli, denominati A e a. Sappiamo che l'allele A (l'allele favorevole al mimetismo) è dominante su a, ma non completamente. Poiché il genotipo dell'animale è dovuto alla combinazione di 2 alleli, gli individui della popolazione possono avere al massimo variazioni di genotipo:
· Genotipo AA: omozigote in cui compare solo la caratteristica dell'allele favorevole al mimetismo. L'animale è in grado di mimetizzarsi perfettamente grazie al colore del suo mantello. Ha probabilità di sopravvivenza massima.
· Genotipo Aa: eterozigote che presenta caratteri intermedi. In sopravvivenza sarà avvantaggiato rispetto a aa, ma svantaggiato rispetto a AA. Il colore del mantello farà mimetizzare l'individuo, ma non bene come l'omozigote favorevole. La sopravvivenza sarà inferiore di una qualche proporzione.
· Genotipo aa: omozigote in cui compare solo la caratteristica sfavorevole. Il mantello non sarà mimetico e le possibilità di sopravvivenza dell'individuo saranno minime.
Differenza dalla codominanza:
_ nella codominanza gli alleli di un medesimo gene sono espressi con piena funzionalità contemporaneamente, come accade nel gruppo sanguigno AB, in cui sono espressi efficacemente entrambi i geni per gli antigeni A e B.
_ nella dominanza incompleta, invece, accade non solo che nell'eterozigote, come avviene anche nella dominanza completa, l'allele mutato è o amorfo, cioè inattivo, o ipomorfo, cioè con attività qualitativamente normale ma quantitativamente ridotta, ma contemporaneamente si osserva anche e soprattutto un'aploinsufficienza della copia selvatica del gene, cioè l'incapacità di una sola copia selvatica del gene per cellula a garantirne una funzione normale, perché per un fenotipo penetrante è richiesto un dosaggio molecolare del prodotto genico più elevato che in eterozigosi.
A
|
a
| |
A
|
AA
|
Aa
|
a
|
Aa
|
aa
|
La codominanza è un evento genetico che si manifesta negli individui diploidi o poliploidi e si riscontra quando due alleli si manifestano entrambi in modo completo, e quindi entrambi sono riconoscibili negli eterozigoti, dal punto di vista del fenotipo.
Un individuo diploide come il Boa Constrictor è dotato di un doppio assetto cromosomico, i due cromosomi di una stessa coppia cromosomica porteranno due alleli (uno per ogni cromosoma) per uno stesso gene.
Gli alleli possono essere uguali e dominanti (omozigote dominante), uguali e recessivi (omozigote recessivo) o diversi (eterozigote).
Nell'eterozigote un allele (dominante) può prevalere sull'altro (recessivo) ed il fenotipo è determinato solo da un allele (quello dominante).
In alcuni casi la dominanza può non essere completa ed in altri casi si può manifestare la codominanza: entrambi gli alleli si manifestano nel fenotipo.
________________________________________________________________________R.R.__Gli alleli possono essere uguali e dominanti (omozigote dominante), uguali e recessivi (omozigote recessivo) o diversi (eterozigote).
Nell'eterozigote un allele (dominante) può prevalere sull'altro (recessivo) ed il fenotipo è determinato solo da un allele (quello dominante).
In alcuni casi la dominanza può non essere completa ed in altri casi si può manifestare la codominanza: entrambi gli alleli si manifestano nel fenotipo.
Il BOA PASTEL
(Sotto: Bci Red Pastel eccellente)

Il pastel nei Boa si ottiene lavorando sull'eredità quantitativa ed è un risultato poligenico, dovuto dall’interazione tra diversi fattori e geni che determinano la pigmentazione, come ad esempio la melanina(pigmento nero) e l'eritrina(pigmento rosso) in diversi gradi, ossia dalla loro totale assenza(fortemente chiaro o appena rosata), fino alla totale saturazione(arancione / rosso carico).
(Sopra: Bci Red Pastel eccellente)
(Sopra: Bci HiperPastel = doppio Pastel eccellente x doppio Pastel eccellente)
I Boa presentati in queste foto sono un'eccellenza selettiva ottenuta prelevando da molteplici cucciolate gli esemplari dalle caratteristiche cromatiche di spicco più estreme ricombinate assieme per più generazioni.
(Sopra: Bci Pastel Ivory)
Per via del loro magnifico patrimonio genetico i veri Pastel accoppiati con altri Boa comuni o morph sono in grado di trasmettere le loro importanti caratteristiche di colorazione e pulizia alla progene diminuendo le antiestetiche puntinature che ne "sporcherebbero" il fondo, migliorandone così l'aspetto.
Solo i ceppi Pastel eccellenti infatti possono essere utilizzati con successo per migliorare e valorizzare ulteriori selezioni genetiche di Boa morph,..
.. vedi come esempio il Pastel Ivory (foto sopra) utilizzato con il morph Anery (foto sotto).
(Sotto: Bci Anery Pastel Ivory)

_____________________________________________________________________R.R.__
BIOGRAFIE:
JOHANN GREGOR MENDEL (1822-1884 )
 IL PADRE DELLA GENETICA
IL PADRE DELLA GENETICA
1) Legge di Segregazione: ciascun carattere ereditato è definito da una coppia di geni. I geni dei genitori vengono suddivisi casualmente nelle cellule sessuali in modo tale che esse contengano solo uno dei due geni della coppia. Quindi, quando le cellule sessuali si uniscono nella fecondazione, la progenie eredita un solo allele da ciascun genitore.
2) Legge dell'Assortimento Indipendente: geni relativi a caratteri differenti si distribusicono nelle cellule sessuali indipendentemente l'uno dall'altro in modo tale che l'ereditarietà di un carattere non dipende dall'ereditarietà di un altro.
3) Legge di Dominanza: un organismo che possiede più forme alternative di uno stesso gene mostrerà la forma dominante.
Gli esperimenti genetici portati avanti da Mendel sulle piante di pisello lo hanno occupato per otto anni (1856-1863) ed egli ha pubblicato le sue conclusione nel 1865. in questo arco di tempo, Mendel ha coltivato oltre 10000 piante di pisello, registrando il numero e il tipo della progenie. Le ricerche di Mendel e le sue Leggi di Ereditarietà non sono state apprezzate nella sua epoca.È stato solo dopo il 1900, successivamente alla riscoperta delle sue Leggi, che sono stati compresi i suoi risultati sperimentali.
 REGINALD CRUNDALL PUNNETT (1875-1967)
REGINALD CRUNDALL PUNNETT (1875-1967)Andò alla Cambridge University come studente di medicina, Punnett ma ottenne laurea in zoologia nel 1898. Dopo la laurea continuò come ricercatore a Cambridge. Egli lavorò sulla morfologia dei Nemertini (vermi piatti). A Punnett sono state dedicate due specie di vermi marini: Cerbratulus punnetti, Punnettia splendia.
A Cambridge Punnett si interessò alle metodologie sperimentali, e scrisse a William Bateson che stava conducendo esperimenti mendeliani su piante e animali. Questo produsse una collaborazione scientifica che contribuì ad avviare la “genetica” a Cambridge. Bateson e Punnett pubblicarono il primo saggio sull’associazione dei geni nel pisello dolce e Punnett ideò il “quadrato di Punnett” per descrivere il numero e la varietà di combinazioni geniche.
Punnett ebbe un ruolo nel collegare i rapporti mendeliani con la statistica. Nel 1908 Punnett fu convocato ad una conferenza, per spiegare perché i fenotipi recessivi continuassero a perdurare – se gli occhi marroni erano dominanti, allora perché l’intero paese non era diventato ad occhi-marroni? Punnett non poté rispondere alla domanda con sua completa soddisfazione. Egli, a sua volta, rivolse la domanda al suo amico e matematico G. H. Hardy. Da questa conversazione nacque la legge di Hardy-Weinberg che calcola come l’ereditarietà genetica agisca su una popolazione.
Nel 1912, quando William Bateson decise di non tornare a Cambridge, Punnett divenne il primo professore della cattedra Arthur Balfour di genetica all’università. Egli lavorò sulla genetica del pisello dolce, del mais e del pollame, sviluppando molte razze. Egli impiegò anche l’associazione genica come mezzo per determinare il sesso nei pulcini. Punnett continuò i suoi esperimenti persino dopo il suo ritiro nel 1940.
Punnett fu un uomo quieto, tollerante e colto che eccelse in tutti gli sport che comportavano l’uso di una piccola e veloce palla. A ottant’anni era ancora un membro attivo del Savile Club di Londra dove giocava a biliardo. Morì nella sua casa nel Somerset, in Inghilterra all’età di 92 anni.
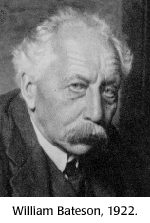
Intorno al 1897, Bateson iniziò a condurre alcuni di questi esperimenti sul pollame e le farfalle. Quando lesse gli scritti di De Vries e di Mendel riconobbe l’importanza della “Legge di Mendel” specialmente in funzione dei propri esperimenti. Dal 1902, Bateson tradusse i lavori di Mendel in inglese e fu un acceso sostenitore delle leggi mendeliane sull’ereditarietà. A Bateson fu attribuito il merito di l’aver coniato i termini “genetica”, “allelomorfo” (successivamente abbreviato in allele), “zigote”, “eterozigote”, e “omozigote”. Nel 1908 come professore di biologia a Cambridge, Bateson contribuì a fondare la Cambridge School of Genetics.
Bateson lasciò Cambridge nel 1910 per accettare l’incarico di direttore della John Innes Horticultural Institute a Merton. Egli continuò ad avere contatti con Cambridge collaborando con R. C. Punnett ad esperimenti di genetica e pubblicazioni. Bateson e Punnett fondarono insieme il Journal of Genetics nel 1910.
Bateson ed i suoi lavori influenzarono gli altro biologi e scienziati come ad esempio: Archibald Garrod, Thomas Hunt Morgan, e Charles Davenport. Bateson ebbe una forte ed agguerrita personalità, ben adattata al ruolo che egli stesso si attribuì come avvocato di Mendel. tuttavia Bateson fu riluttante a credere nella teoria cromosomica dell’ereditarietà. Egli fu un oppositore verbale di questa teoria fino al 1922, quando dopo la visita al Thomas Hunt Morgan's fly lab egli accettò pubblicamente i cromosomi e il loro ruolo nell’ereditarietà
LINNEO


 Carl Nilsson Linnaeus, divenuto Carl von Linné in seguito all'acquisizione di un titolo nobiliare e noto ai più semplicemente come Linneo (dalla forma latinizzata del nome, Carolus Linnaeus) (Råshult, 23 maggio 1707 – Uppsala, 10 gennaio 1778), è stato un medico e naturalista svedese, considerato il padre della moderna classificazione scientifica degli organismi viventi.
Carl Nilsson Linnaeus, divenuto Carl von Linné in seguito all'acquisizione di un titolo nobiliare e noto ai più semplicemente come Linneo (dalla forma latinizzata del nome, Carolus Linnaeus) (Råshult, 23 maggio 1707 – Uppsala, 10 gennaio 1778), è stato un medico e naturalista svedese, considerato il padre della moderna classificazione scientifica degli organismi viventi.La lettera L., posta spesso a seguire delle indicazioni di nomenclatura binomiale nei cataloghi di specie, identifica il cognome dello scienziato.,Il merito maggiore dello svedese fu la definizione e l'introduzione nel 1735 della nomenclatura binomiale, basata sul modello aristotelico di definizione mediante genere prossimo e differenza specifica, nel sistema di classificazione delle piante e degli animali.[2] Con questo metodo tassonomico (concepito poco più di un secolo prima da Bauhin) a ciascun organismo sono attribuiti due nomi (di origine latina): il primo si riferisce al Genere di appartenenza dell'organismo stesso ed è uguale per tutte le specie che condividono alcuni caratteri principali (nomen genericum); il secondo termine, che è spesso descrittivo, designa la Specie propriamente detta (nome triviale o nome specifico).



Buon allevamento!